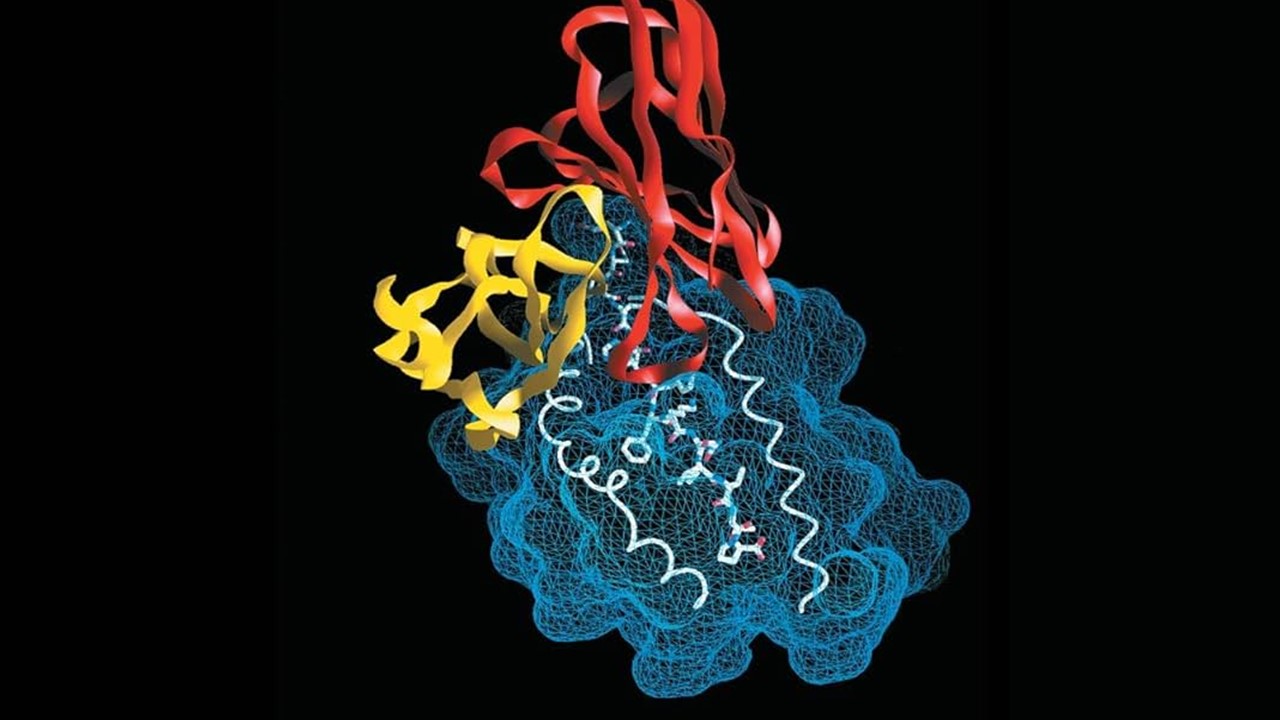
The Evolutionary Tapestry of Photosystem I
Photosystem I (PSI), a cornerstone of oxygenic photosynthesis, has undergone remarkable structural evolution across species. In cyanobacteria, PSI exists as a trimer, a configuration optimized for energy capture in early Earth’s low-light environments. This trimeric architecture, preserved in thermophilic species like Thermosynechococcus elongatus (Te), contrasts with the monomeric form found in plants, reflecting adaptations to divergent ecological niches. The retention of trimeric PSI in cyanobacteria offers a window into primordial photosynthetic machinery, where lipid-protein interactions likely played a pivotal role in stabilizing these complexes within galactolipid-rich thylakoid membranes.
Recent studies suggest that PSI’s oligomeric state is influenced by its lipid microenvironment. Cryo-EM analyses reveal detergent-induced structural perturbations in PSI trimers, hinting at lipids’ role in maintaining integrity. The Jordan et al. crystal structure identified four tightly bound lipids near PSI’s core, including one coordinating a chlorophyll molecule, underscoring lipids as functional cofactors rather than passive bystanders. These findings challenge the notion of lipids as mere structural scaffolds, positioning them as active participants in photosynthetic efficiency.
The dynamic interplay between PSI and its lipid annulus raises questions about how extraction methods alter native configurations. Traditional detergent-based isolation strips lipids, potentially distorting protein structure and function. In contrast, emerging techniques like styrene-maleic acid lipid particles (SMALPs) preserve native lipid environments, offering unprecedented insights into PSI’s natural state. This dichotomy sets the stage for exploring how lipid retention influences PSI’s photochemical prowess.
SMALP Technology: Preserving Nature’s Blueprint
Styrene-maleic acid copolymers (SMAs) have revolutionized membrane protein research by enabling the extraction of protein-lipid complexes without disruptive detergents. SMALPs encapsulate proteins within a native lipid bilayer, encircled by a copolymer belt. For PSI, this approach retains over 1,200 lipids per trimer—a stark contrast to the mere handful surviving detergent isolation. The SMA 1440 copolymer, with its unique esterified maleic acid groups, exhibits enhanced affinity for cyanobacterial thylakoids, likely due to interactions with anionic lipids like sulfolipids.
Neutron reflectometry studies reveal SMA 1440’s deep insertion into lipid monolayers, particularly those rich in sulfolipids. This selective partitioning suggests SMALP formation is guided by lipid composition, favoring regions enriched in specific anionic species. The resulting PSI-SMALP particles, the largest yet reported, preserve not only lipids but also chlorophyll orientation, as evidenced by a 3 nm red shift in fluorescence—a hallmark of native antennae configuration.
The thermostability of Te PSI further enhances SMALP utility. Growth in deuterated media enables isotopic labeling, reducing incoherent neutron scattering and allowing precise contrast matching. This capability, paired with SMA 1440’s defined belt structure (~1 nm thickness), positions SMALPs as indispensable tools for probing membrane protein dynamics in near-native conditions.
Small-Angle Neutron Scattering: Mapping Molecular Landscapes
Small-angle neutron scattering (SANS) offers unparalleled resolution in analyzing soft matter like PSI-SMALPs. By exploiting deuterium’s distinct scattering properties, contrast variation techniques isolate contributions from protein, lipid, and copolymer components. For PSI-SMALP, SANS revealed a discoidal particle ~354 Å in diameter, with a lipid annulus accounting for ~30% of its size compared to detergent-isolated PSI.
The radius of gyration (Rg) and pairwise distance distribution (P(R)) analyses confirmed PSI-SMALP’s monodispersity and elongated shape. At full contrast, the maximum dimension (DMAX) reached ~354 Å, shrinking to ~344 Å when the SMA belt was contrast-matched. This subtle difference underscores the copolymer’s tight adherence, minimizing protrusion beyond the lipid bilayer. In contrast, detergent-solubilized PSI exhibited a 29 Å reduction at its contrast match point, reflecting the loss of its native lipid shell.
These dimensional insights align with lipidomic data: the retained lipid annulus in SMALPs spans ~450 nm², accommodating ~1,300 lipids—40-fold more than detergent-isolated PSI. This lipid reservoir likely buffers against structural fluctuations, maintaining PSI’s functional geometry. The correlation between SANS-derived dimensions and experimental lipid counts validates SMALP’s fidelity in preserving membrane microenvironments.
Lipidomics: A Surprising Mosaic of Molecular Species
Lipidomic profiling of PSI-SMALP unveiled a striking divergence from bulk thylakoid composition. Sulfolipid SQDG dominated, constituting over half the lipid content—a stark enrichment compared to its modest presence in native membranes. This anionic lipid’s prevalence near PSI suggests electrostatic interactions with positively charged protein residues, potentially stabilizing the trimeric interface or optimizing chlorophyll orientation.
Detergent-isolated PSI, by contrast, retained negligible SQDG, highlighting detergent-induced lipid stripping. SMALP’s lipid profile also showed heightened saturation, with balanced 32- and 34-carbon acyl chains. Such saturation may enhance membrane rigidity, countering thermal motion in Te’s high-temperature habitat. Notably, DGDG and MGDG—galactolipids critical for membrane curvature—were abundant, though their ratios shifted significantly from bulk membranes.
The functional implications are profound. SQDG’s enrichment correlates with accelerated photochemical kinetics in PSI-SMALP, including ultrafast charge separation observed via femtosecond spectroscopy. This lipid’s negative charge might facilitate electron transfer or stabilize transient states during photooxidation. Conversely, detergent-isolated PSI, stripped of SQDG, exhibits sluggish reduction kinetics, underscoring lipids’ role in optimizing reaction center dynamics.
Chlorophyll Dynamics: A Sensor of Structural Integrity
Flash photolysis studies exposed stark differences in PSI-SMALP and detergent-isolated PSI. SMALP-retained PSI demonstrated a sixfold faster P700+ reduction rate, attributed to preserved chlorophyll networks and lipid-mediated electron pathways. The antennae chlorophyll count in SMALPs matched theoretical maxima (112 per monomer), whereas detergent isolation caused a 16% loss, aligning with discrepancies in prior crystal structures.
The retained chlorophylls in SMALPs likely maintain optimal energy transfer distances, minimizing exciton loss. Detergent-induced chlorophyll displacement disrupts this network, blunting photochemical efficiency. These findings underscore lipids’ role in anchoring pigments—a function compromised by traditional isolation methods but preserved in SMALPs.
Moreover, the linear correlation between chlorophyll content and PSI concentration in SMALPs validates BCA protein assays, previously questioned due to detergent-pigment interactions. This consistency reinforces SMALP’s reliability for quantitative studies, bridging structural and functional analyses.
Biotechnological and Evolutionary Implications
The PSI-SMALP model reshapes our understanding of photosynthetic machinery. The lipid annulus, enriched in SQDG, may represent a specialized membrane domain optimized for PSI trimerization and function. Such domains, preserved through evolution in cyanobacteria, could inform strategies to engineer artificial photosynthetic systems with enhanced efficiency.
SMALP technology’s ability to isolate intact complexes opens avenues for studying other membrane proteins in native environments. The preferential extraction of PSI over PSII or cytochrome complexes hints at lipid-driven selectivity, a feature exploitable for targeted protein purification. Furthermore, Te’s resilience in deuterated media paves the way for neutron crystallography, offering atomic-level insights into lipid-protein interactions.
Ultimately, this work bridges structural biology and ecology, illustrating how lipid environments underpin the evolutionary success of photosynthetic organisms. By preserving these interactions, SMALPs not only illuminate nature’s design principles but also inspire biomimetic innovations in energy conversion and nanotechnology.
Photosystem I: Redefined
This synthesis of neutron scattering, lipidomics, and photochemical analyses redefines PSI as a lipid-protein supercomplex, where every molecular component is a deliberate player in the photosynthetic symphony. The lessons learned here transcend cyanobacteria, offering a blueprint for exploring membrane biology across the tree of life.
Study DOI: https://doi.org/10.1016/j.bbabio.2022.148596
Engr. Dex Marco Tiu Guibelondo, B.Sc. Pharm, R.Ph., B.Sc. CpE
Editor-in-Chief, PharmaFEATURES

Subscribe
to get our
LATEST NEWS




Related Posts
Medicinal Chemistry & Pharmacology
Aerogel Pharmaceutics Reimagined: How Chitosan-Based Aerogels and Hybrid Computational Models Are Reshaping Nasal Drug Delivery Systems
Simulating with precision and formulating with insight, the future of pharmacology becomes not just predictive but programmable, one cell at a time.
Medicinal Chemistry & Pharmacology
Coprocessed for Compression: Reengineering Metformin Hydrochloride with Hydroxypropyl Cellulose via Coprecipitation for Direct Compression Enhancement
In manufacturing, minimizing granulation lines, drying tunnels, and multiple milling stages reduces equipment costs, process footprint, and energy consumption.

Medicinal Chemistry & Pharmacology
Decoding Molecular Libraries: Error-Resilient Sequencing Analysis and Multidimensional Pattern Recognition
tagFinder exemplifies the convergence of computational innovation and chemical biology, offering a robust framework to navigate the complexities of DNA-encoded science